 |
|
| 研究紹介 | メンバー | 研究成果 | 写真 | ブログ | セミナー | リンク | |
トップページ > 研究紹介 > ホヤの変態メカニズム
ホヤの変態メカニズム
ホヤは、幼生の時期には写真のような「オタマジャクシ」の形態をとり、尾部を振って活発に泳ぎ回ります。遊泳幼生はやがて岩などに固着し、尾部を失うなど変態を始め、成体の形に変化します。ホヤの成体は、活発に運動する幼生とは異なりほとんど動き回らない固着生活を送り、海水中のプランクトンや有機物を濾し取って食べる濾過摂食を行います。カタユウレイボヤでは、変態イベントは脊椎動物や昆虫のゆっくり進行する変態とは大きく異なり、およそ2日程度で完了する非常にダイナミックなものです。この素早い変態は、変態のメカニズムを研究する上で非常に有利になります。
 |
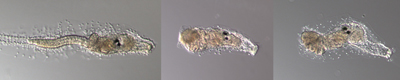 |
| カタユウレイボヤの幼生(上)と変態中のホヤ(下) 尾部が吸収し、付着突起の形態も変化する。下の段の一番右が変態直後の幼若体。幼生から幼若体へとダイナミックに形態が変化する。 |
ホヤの成体は一度変態するとほとんど動けません。つまり、どこで変態するかを決めることはホヤにとって非常に重要な問題になります。例えば他の個体と遠く離れたところで一匹だけ変態すると、生殖に問題が生じます。実際のところ、ホヤの変態は複雑な制御を受けていることが分かってきていますが、その全体像はほとんど分かっておらず、その制御機構を解明することがホヤという動物の理解に重要になります。また、ホヤは脊椎動物に最も近い動物群であり、一生涯遊泳生活を送るタイプの祖先型動物から進化してきたであろうことが示されています。つまりホヤは脊椎動物と分かれてから、変態するように進化していったと考えられています。そのホヤは脊椎動物と共通の遺伝子セットを持っていることがゲノムプロジェクトから分かっていますが、脊椎動物と共通の遺伝子セットを使いながら、ホヤがどのような過程で変態し固着生活を送るようになったのか、その進化的なプロセスも興味のあるところです。これらの謎を明らかにするためには、ホヤの変態がどのような遺伝子により制御されているのか、そのメカニズムを解明することが重要になります。このような観点から、我々の研究グループでは、このホヤの変態のメカニズムに着目して研究を進めています。
ホヤの変態はおよそ10個のイベントからなることが示されています。代表的なイベントは「固着」「尾部の吸収」「体軸の回転」「成体組織の成長」です。これらの変態イベントは、固着して初めて開始されます。幼生はふ化後数時間すると変態する能力を獲得しますが、その変態能力の獲得後も、遊泳している間は変態を始めません。考えてみるとこれは当たり前のことです。遊泳している間に変態してしまうと、尾部が無くなるのでそれ以上遊泳出来なくなるため、海底に落ちてしまうだけですから。幼生の体幹部前方には「付着突起」と呼ばれる突起物があります。この部分は付着するのに必要な粘着物質を出しています。そのためホヤの幼生は付着突起で固着します。また、付着突起は神経系の組織であり、脳と連結しています。固着の刺激はこの付着突起の神経の興奮という形で、幼生の後方に伝えられ、他の変態のイベントを開始させるのだと考えられています。このようなことから、基本的には固着することが刺激となり、変態が開始されるのだと考えられています。
突然変異体から変態メカニズムを解明する
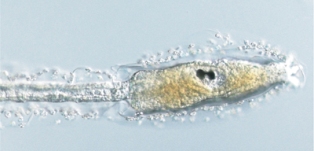
ホヤの変態のメカニズムを解明するのには、その変態イベントに異常を生じる突然変異体の単離が有効な手法です。我々の開発したトランスポゾンを用いた遺伝子破壊方法(リンク)により、変態イベントに異常を示す突然変異体を作製しています。 上の写真がその一例です。先ほど述べたように、ホヤの幼生は固着して初めて変態を開始します。しかしこの突然変異体では、幼生が泳いでいる、つまり固着していないにも関わらず、幼生が変態イベントの一部(体軸の回転など)を始めてしまいます。ただ、この突然変異体で生じる変態イベントは一部に限られ、尾部の吸収などの変態イベントは生じません。この突然変異体は、体幹部(ホヤ幼生の前方部)は変態直後の個体(幼若体と呼びます)と同じような形をしていますが、完全な尾部を持ち遊泳し続けることから、「泳ぐ幼若体」という意味の swimming juvenile と名付けられました。この突然変異体から、ホヤの幼生は固着すると変態を開始するだけでなく、遊泳している時には変態しないように、変態イベントを抑制する仕組みを持つということが分かります。この抑制に関わる遺伝子が変異によりつぶれた結果、泳ぎながら変態を始めてしまう変異体が生じたのです。
 |
 |
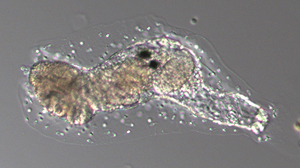 |
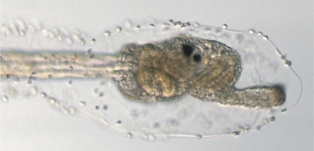
| 正常な幼生 | swimming juvinile変異体の幼生 | 変態後の正常な幼生 |
| 変異体の幼生の頭は、次第に変態後の個体の形態に似てくる。 | ||
swimming juvenile 突然変異体では、セルロース合成酵素をコードする遺伝子が Minos トランスポゾンにより破壊されていました。つまり、セルロース合成酵素が働かなくなり、そのことが原因でホヤの変態がおかしくなったと考えられます(ホヤのセルロース合成酵素に関してはこちらのリンクを参考にして下さい)。つまりセルロース合成酵素は、幼生の時期に変態イベントの一部が進行しないように抑制している機構に関与しているということが推察されます。カタユウレイボヤのセルロース合成酵素には、セルロースを作り被のうを形成するだけでなく、ホヤの変態イベントを正常に進行させる役割を持っているということです。
セルロース合成酵素がどのような仕組みでホヤの変態現象を制御しているのか、その仕組みはまだ解明されていません。またセルロース合成酵素により影響を受けない変態イベントも存在しています。ホヤの変態がどのような仕組みで生じているのか、その全体像はまだまだ謎に包まれています。現在我々の研究グループでは、 swimming juvenile の他にも変態現象に異常を生じる突然変異体を単離しています。これらの突然変異体で異常になる遺伝子を特定することにより、ホヤ変態を制御する遺伝子が同定出来ます。また、変態時期に発現する遺伝子をゲノミクスやプロテオミクスの手法を用いて同定するプロジェクトを進めています。このような手法を駆使することにより、ホヤ変態を制御する遺伝子カスケードの全体像を明らかにし、ホヤの変態の謎を解明することを目指しています。